Корзина пуста

rhIL-4, интерлейкин 4 человека, рекомбинантный белок

Синонимы: Interleukin 4, IL-4, B cell differentiation factor, BCDF, B cell stimulatory factor 1, BSF1, B cell growth factor 1.
Источник: Клеточная линия CHO, продуцирующая rhIL-4.
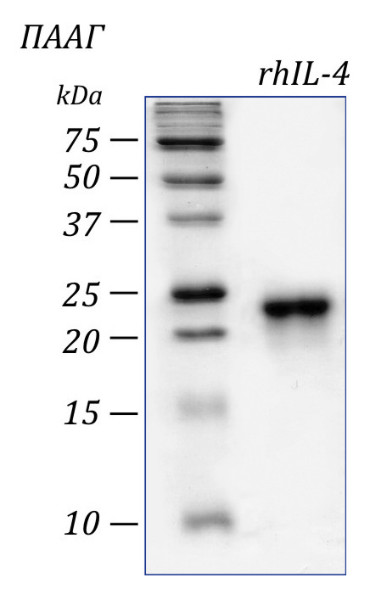
Чистота: >98% в соответствии с электрофорезом в ПААГ с последующим окрашиванием Coomassie Brilliant Blue.
Уровень эндотоксина: <1.0 EU на 1 мкг белка, LAL-тест.
Молекулярный вес: 24 кДа в редуцирующих условиях в ПААГ.
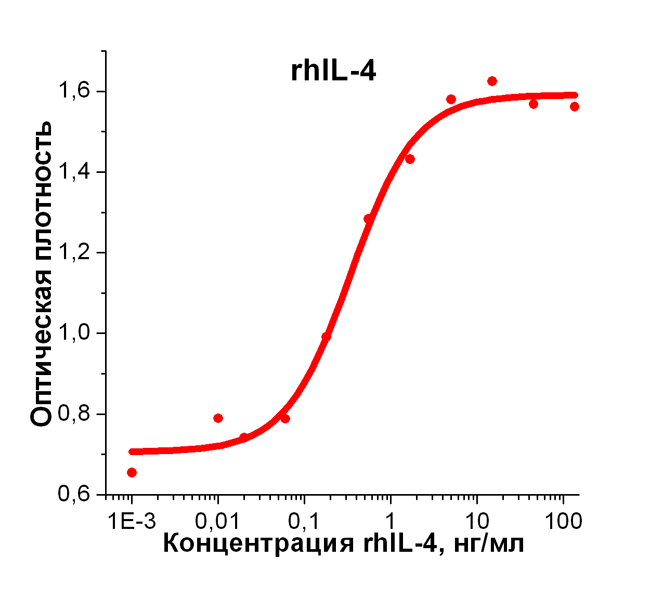
Биологическая активность: Рекомбинантный белок, rhIL-4 человека, стимулирует пролиферацию клеток линии эритролейкоза человека (TF-1). ED50 для данного эффекта обычно 0,4 - 1 нг/мл. Оптимальная концентрация для индивидуального применения определятся пользователем.
Форма
Лиофильно высушен из фосфатного буферного раствора (PBS), содержащего 0,05% Tween20, pH 7.0, профильтрованного через фильтр с диаметром пор 0,22мкм.
Не содержит вспомогательных белков.
Разведение
Центрифугировать флакон при 1000g, 3 мин. Добавить стерильный фосфатный буферный раствор (PBS) до конечной концентрации 0,1-1 мкг/мкл. Оставить на 20-30 мин при комнатной температуре, затем центрифугировать при 1000g в течение 1 мин, и мягко ресуспендировать. Для приготовления рабочих растворов можно использовать буфер на водной основе или культуральную среду. Добавление вспомогательных белков (BSA или FBS) не является необходимым.
Условия транспортировки
Перевозить при комнатной температуре.
Стабильность и Условия хранения
- 24 месяца, хранение невскрытой упаковки, при температуре от -20 до -70°C.
- 1 месяц, разведенный в стерильных условиях, при температуре от 2 до 8°C.
- 6 месяцев, разведенный в стерильных условиях, при температуре от -20 до -70°C.
Не рекомендуются повторные циклы замораживания-оттаивания раствора рекомбинантного белка.
rhIL-4 (#PSG040) был использован в следующей работе
Glycan recognition by human blood mononuclear cells with an emphasis on dendritic cells
Eugenia M. Rapoport, Sergey V. Khaidukov, Andrey M. Gaponov, Galina V. Pazynina, Svetlana V. Tsygankova, Ivan M. Ryzhov, Ivan M. Belyanchikov, Panagiota Milona, Nicolai V. Bovin, Kenneth C. McCullough.
Glycoconjugate Journal volume 35, pages191–203(2018); https://doi.org/10.1007/s10719-017-9811-6
| Описание | |
| Дополнительное описание и цитирование | Интерлейкин-4 (ИЛ-4) (IL-4, от англ.Interleukin-4) - плейотропный цитокин, регулирующий клеточный и гуморальный иммунный ответ, участвующий в процессах пролиферации, дифференцировки и активации клеток различного типа. IL-4 экспрессируется активированными Т-лимфоцитами, тучными клетками, дендритными клетками, NKT-клетками, базофилами, эозинофилами. Эпителий и опухолевые клетки являются источниками IL-4 в тканях. IL-4 проявляет свою функциональную биологическую активность, связываясь со специализированными рецепторами на поверхности клеток мишеней. Описано три типа рецепторных комплексов для IL-4. Тип I является гетеродимером и состоит из лиганд-связывающей IL-4Rα цепи и g -цепи (общей субъединицы рецепторов IL-2, IL-7, IL-9,IL-15, IL-21). Рецептор типа II представляет собой гомодимерный комплекс IL-4Rα или гетеродимерный комплекс IL-4Rα и IL-13Rα1 цепей, последний является общим для IL-4 и IL-13. IL-4R-I экспрессируется на гемопоэтических клетках лимфоидного (Т- и В-лимфоциты) и миелоидного (моноциты, макрофаги и фибробласты) происхождения. Связывание IL-4 с IL-4Rα индуцирует гетеродимеризацию с g-цепью и последующую активацию тирозин киназ Янус семейства JAK1 и JAK3. IL-4R-II экспрессируется на гемопоэтических клетках миелоидного происхождения и негемопоэтических клетках (гладкомышечные и эпителиальные клетки), а так же на поверхности опухолевых клеток. Связывание с IL-4R-II ведет к активации Янус-киназ - JAK1, JAK2 и TYK2. В результате проведения сигнала обоими типами IL-4 рецепторов происходит фосфорилирование цитоплазматического транскрипционного фактора STAT6. STAT6 является ключевым фактором регуляции экспрессии генов вовлеченных в развитие широкого спектра эффекторных функций IL-4 в различных типах клеток. В В-лимфоцитах запуск каскада фосфорилирования по пути IL-4R/STAT6 индуцирует экспрессию генов, вовлеченных в В-клеточную дифференцировку и переключение синтеза изотипов иммуноглобулинов на IgE, IgG1 и IgА. В Т-клетках фосфорилирование STAT6 активирует экспрессию генов E4BP4 и GFI-1, вовлеченных в регуляцию пролиферации и повышению жизнеспособности Т-лимфоцитов и генов GATA3 и CRTH2 определяющих дифференцировку Т-лимфоцитов в Th2-клетки. IL-4/STAT6 сигнальный каскад играет существенную роль в стабилизации Th2 фенотипа, и ингибироанию дифференцировки по Th1 и Th17-пути. Активация Th2-лимфоцитов и синтез иммуноглобулинов приводит к активации тучных клеток, базофилов, эозинофилов и развитию гуморального иммунного ответа. Нарушение регуляции Т-клеточного отклика лежит в основе развития хронического воспаления, приводит к развитию аллергических реакций, атопического дерматита (АД), астмы. Хотя патогенез АД до конца не изучен, но показано, что избыточная экспрессия IL-4 Т-лимфоцитами и дисрегуляция экспрессии генов кератиноцитов, лежат в основе развития АД. IL-4, активируя сигнальный путь IL-4Ra/STAT6, стимулирует повышение продукции хемокинов (CCL8, CCL24, CCL26 и др.), про-воспалительных (IL-1a, IL-19, IL-20, IL-25 и др.) и про-ангиогенных факторов, снижает уровень антимикробных пептидов и факторов, ответственных за реализацию барьерных функций кожи и, в результате, приводит к развитию АД. Формирование острого аллергического воспаления в дыхательных путях в значительной степени проходит по пути IL-4/IL-13Rα1/STAT6 и приводит к инфильтрации области воспаления эозинофилами, макрофагами и дендритными клетками, усиливает пролиферацию и активацию фибробластов. Стимуляция эпителиальных клеток дыхательных путей приводит к увеличению выработки слизи, активация гладкомышечных клеток к гиперчувствительности дыхательных путей. Второй сигнальный путь, активируемый IL-4, ассоциирован с субстратом-2 рецептора инсулина (insulin receptor substrate 2, IRS-2). IL-4 индуцирует JAK1 и JAK3 киназную активность, что приводит к фосфорилированию тирозинового остатка Tyr497 трансмембранного домена IL-4Rα, который является док-сайтом для сигнальной адаптороной молекулы IRS-2. Мишенями для IRS-белков являются регуляторная субъединица p85 фосфатидилинозитол-3-киназы (PI3K) и адаптор Grb-2. Активация данных сигнальных каскадов непосредственно связана с регуляцией пролиферации, устойчивости к апоптозу и индукцией генов, ассоциированных с «альтернативным» путем активации макрофагов в М2-клетки и ингибированием классической активации макрофагов в M1. Увеличение M2 макрофагов в сочетании с секрецией ими IL-10 и TGF-β приводят к уменьшению патологического воспаления. Секреция аргиназы, пролина, TGF-β активированными М2 клетками связана с заживления ран. Оказывая свое действие на пролиферативную способность и функциональную активность В- и Т-лимфоцитов, тучных клеток, эндотелиальных и эпидермальных клеток, запуская «альтернативную активацию макрофагов», IL-4 является доминантным цитокином в поддержании баланса между формированием иммунного ответа организма и развитием аллергического воспаления. Использованная литература:
|